Description
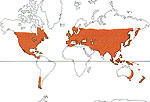
Distribution
Les Fagaceae sont une famille importante d'arbres au bois dur, et plus rarement d'arbustes. Elles représentent une biomasse considérable, presque comparable à celle des Conifères.
Ce sont des espèces qui dominent les forets de feuillus qui couvrent, ou couvraient, de vastes régions de moyennes latitude de l'hémisphère nord, et, dans une moindre mesure, des régions de l'hémisphère sud.
Les hêtres (
Fagus), les chênes (
Quercus) et les châtaigniers (
Castanea) dominent dans les forets à feuillage caduc et mixtes d'Amérique du nord et d'Eurasie. Dans les forets du même type, le long du sud de la cordillère des Andes, c'est le hêtre austral (
Nothofagus) qui domine.
Les chênes à feuillage persistant peuplent les forets autour du golfe du Mexique, dans les régions sud de la Chine et du Japon, et les hêtres austraux à feuille persistantes dominent les forets du sud de la Nouvelle Zélande. Dans le sud de l'Asie, la structure de la foret mixte de montagne est déterminée par des genres à feuilles persistantes de la famille, particulièrement les chênes et les hêtres austraux.
Appareil végétatif
Les Fagaceae sont des arbres, plus rarement des arbustes, au feuillage caduc ou persistant. Leurs feuilles sont alternes, parfois pseudo-verticillées (Cyclobalanopsis), simples, entières ou pennatilobées et pourvues généralement de stipules caduques. Les racines forment fréquemment des ectomycorhizes avec des champignons à hyphes.
Anatomie
L'anatomie des Fagaceae montre un periderme superficiel et un pericycle hétérogène. Des poils tecteurs unicellulaires, isolés ou en bouquets, sont aussi présents. Les poils sécréteurs sont très rares. Le liber secondaire des tiges âgées renferme des fibres ou des cellules scléreuses.
Reproduction
Les fleurs sont toujours unisexuées, et il y a généralement monécie, plus rarement dioecie. Leurs inflorescences sont très variées : elles sont formées de cymes bipares complètes ou réduites, parfois à une fleur, isolées ou groupées en chatons ou en capitules ; les inflorescences femelles comptent de 1-5 fleurs. Chez Nothofagus, les fleurs mâles sont solitaires. Chez Castanea, un caractère ancestral consiste en la présence de fleurs femelles à la base des inflorescences mâles.
Les fleurs sont toutes apétales. Les mâles possédent 3-12 sépales scaliformes, libres ou connés, et 3-40 étamines libres, aux anthères biloculaires dehiscentes par des fentes longitudinales, et parfois un rudiment de pistil. Les fleurs femelles ont 4-6 sépales rudimentaires, libres ou connés, et un ovaire infère constitué de 3-6 carpelles indépendants, chacun contenant 2 ovules pendants, avec une placentation axille.
L'origine phylogénétique et la valeur morphologique des fleurs des Fagaceae restent controversées.
Les fruits sont des nucules monospermes enclos par 1-5 dans des cupules qui portent sur leur face externe divers appendices, telles les écailles des cupules de chênes, les épines ramifiées des bogues de châtaigniers. Les graines n'ont pas d'albumen, et ont un embeyon droit.
Le pollen et d'autres caractères, comme l'inflorescence très odorante, suggèrent que la pollinisation par les insectes est un caractère ancestral des Fagaceae. Elle est conservée chez de nombreux genres, sauf chez Fagus, Nothofagus et les espèces de Quercus des régions tempérées.
La présence des cupules est le caractère le plus remarquable des Fagaceae. Ces organes entourent soit une fleur isolée, soit une cyme, et comportent ou non des valves. Le cas le plus primitif est celui des cupules composées de trois valves qui entourent une seule fleur et donc un seul fruit. Les cupules cupuliformes (chênes) ou urcéolées, non découpées en valves, résultent d'une évolution plus poussée qui s'est produite indépendamment chez les Castaneoïdeae et les Quercoïdeae. L'origine des cupules est très discutée.
La cupule des Fagaceae est parfois considérée comme étant le resultat de la condensation d'une involucre. Les appendices des cupules ont été assimilés à des feuilles du fait qu'ils présentent souvent une disposition phyllotaxique et qu'ils sont parfois pourvus de faisceaux. Mais ces arguments ont été réfutés par Forman (1966) qui montra que les appendices des cupules sont des émergences, c'est-à-dire des productions superficielles de l'épiderme, et que la cupule elle-même est une structure intercalaire, représentée dans son état primitif par une expansion trilobée du pédicelle à la base de chaque fleur femelle ; la cupule est donc de nature caulinaire. C'est seulement avec la découverte en 1961 du genre Trigonobalanus, limité au nord de Bornéo, aux Célèbes, au nord de la Thaïlande, de la Malaisie et du Sarawak, qu'il a été possible d'infirmer cette hypothèse. Des anomalies, telle la présence de fleurs sur la face externe des cupules de Castanea sativa ou au sommet des valves de Fagus, appuient aussi cette hypothèse. Il est possible que la cupule des Fagaceae représente un chaînon avec les Pteridospermophytes, ancêtres des plantes à fleur. Enfin, Fey et Endress (1983) ont montré que la cupule serait une structure complexe resultant de la soudure, de la condensation et de la lignification d'axes inflorescentiels de niveaux de ramification variés, portant des bractées modifiées en écailles ou en épines.
Enfin, les fruits des Fagaceae ont une capacité de dispersion lente et limitée, et leur pouvoir germinatif diminue rapidement avec le temps. Ces caractères, conjugués avec celui de l'ancienneté de la famille, sont à l'origine de discussion concernant la migration des plantes occasionnée par la dérive des continents, une évidence dans le cas des Nothofagus.
Classification et phylogénie
L'évolution de la famille, bien que très ancienne, s'est faite dans un domaine restreint. Certains genres diffèrent peu entre eux et des caractères semblables existent dans des groupes éloignés, soit qu'ils aient été hérités d'ancêtres communs, soit qu'une évolution parallèle de deux branches phylogénétiques différentes ait abouti à des différenciations semblables.
Des fossiles de Fagaceae ont été trouvés depuis le milieu du Crétacé, soit 90 millions d'années. Apparues sous un climat plus chaud qu'aujourd'hui, à proximité de l'équateur, les Fagaceae ont migré ensuite vers les pôles. Leur ancienneté explique certains traits archaïques : lors de la fécondation, le tube pollinique n'entre pas dans l'ovule par son sommet, ce qui est le cas général, mais latéralement ou même à sa base ; les éléments du bois possèdent encore des perforations de type scalariforme ; et les fleurs sont encore construites sur un type 3 ou 2.
La famille des Fagaceae se divise en 3 sous-familles. Si l'accord est presque général sur leur distinction et leur délimitation, il n'en est pas de même pour les genres. Ce sont les conceptions de Camus (1929-1954) et de Forman (1966) qui sont admises actuellement, alors que certains genres reconnus par Schwarz (1936) et par Hjelmqvist (1948) restent discutés.
Les Fagoïdeae ont des fleurs solitaires ou en grappes plutôt arrondies ; leurs cotylédons sont épigés et foliacés. Elles comptent deux genres : Fagus, les hêtres aux inflorescences mâles longuement pédonculées et aux fleurs nombreuses, avec des styles longs et Nothofagus, les hêtres australs aux inflorescences mâles sessiles ou courtement pédonculées et aux styles courts.
Les Castaneoïdeae ont des fleurs en chatons allongés simples ou composés de cymes bipares ; les cotylédons sont hypogés et épais ; les fleurs mâles ont ordinairement 12 étamines, aux anthères dorsifixes, et possèdent un rudiment de pistil ; les stigmates des fleurs femelles sont petits et ponctiformes. On distingue 4 genres : Chrysolepis de l'ouest des États-Unis, dont la cupule est divisée en valves libres ; Castanopsis du sud de la Chine et de l'Indo-Malaisie à la cupuler aux valves soudées ; Castanea de l'hémisphère Nord, aux feuilles caduques, aux fleurs femelles à 6 ou plus carpelles ; Lithocarpus principalement dans le sud-est asiatique, au cupules sans valves.
Les Quercoïdeae ont des fleurs en chatons allongés simples ou composés de cymes bipares ; les cotylédons sont hypogés et épais ; les fleurs mâles ont ordinairement 6 étamines, aux anthères plus ou moins basifixes, et sont dépourvues de rudiment de pistil ; les stigmates des fleurs femelles sont grands. Elles comptent 2 genres : Quercus, les chênes, avec 600 espèces dont 25 indigènes en Europe, avec des fleurs femelles solitaires, des fruits ronds et une cupule non lobée ; Trigonobalanus, aux fleurs femelles groupées en grappes de 3-7 fleurs, aux fruits à trois angles et à la cupule lobée.
Les deux espèces de Trigonobalanus sont : T. verticillata, avec beaucoup de caractères ancestraux et des feuilles verticillées par trois, de Bornéo et de Célèbes, et T. doichangensis du nord de la Thaïlande.
Intérets
La famille fournit une des plus importantes sources mondiales de bois durs, particulièrement avec le chêne, le hêtre et la châtaignier. Cette ressource conjuguée avec le défrichement dû à l'agriculture a contribué à la destruction de vastes surfaces occupées par la famille.
Malgré leur bois de bonne qualité, les genres tropicaux Castanopsis et Lithocarpus sont peu exploités.
Le bois des Quercus est très dur, reconnaissable à de larges rayons médullaires ; il est utilisé comme bois de construction, ébénisterie, et c'est également un bon combustible. Parfois, l'écorce est utilisée en tannerie. Le gland entre dans la nourriture des porcs, mais sa teneur élevée en tanins restreint son utilisation comme aliment de l'homme en temps de rudes disette. L'écorce du Q. suber est le liège des bouchons. Le Q. ballota donne des glands non astringents qui, torréfies fournissent le café de glands doux. Sous l 'action de piqûres d'insectes, divers chênes produisent des galles d'où on extrait le tanin.
Castanea sativa, du sud de l'Europe, est parmi les châtaigniers l'espèce la plus cultivée, et ses châtaignes sont consommées sous de nombreuses formes.
Données éditées le 27 novembre 2005












