L'inflorescence est caractéristique de la famille, et consiste en une grande spathe souvent pétaloïde, sous-tendant et parfois enveloppant un spadice formé de nombreuses petites fleurs pouvant être bisexuées ou unisexuées.
L'appareil reproducteur est relativement varié. Les spathes, parfois de couleur verdâtre, terne, peuvent former des cornets d'un blanc pur (
Zantedeschia aethiopica) ou d'un rouge éclatant (
Anthurium andreanum). Elles sont caduques ou persistantes, parfois seulement à la base. Elles sont absentes chez quelques genres (
Orontium,
Gymnostachys). Le spadice est parfois très réduit, ne comportant que quelques fleurs (
Ambrosinia) ou, au contraire, gigantesque (plus d'un mètre de long chez certains
Amorphophallus), couvert en totalité (
Acorus, Philodendron) ou partiellement (
Arum) de fleurs.
 Morphologie d'une inflorescence chez Arum maculatum
Morphologie d'une inflorescence chez Arum maculatum
A gauche, l'aspect global, avec un spadice entouré d'une grande bracée, la spathe. Au centre, le détail d'un spadice, avec, de bas en haut, des fleurs femelles, des fleurs mâles, un anneau cilié stérile intervenant lors de la pollinisation entomophile et un long appendice renflé apicalement, stérile lui aussi. A droite, de bas en haut, le détail d'une fleur femelle et d'une fleur mâle, toutes deux de structure extremement simplifiées.
Les fleurs sont toujours très petites et sessiles. Elles sont, soit bisexuées, périanthées ou plus réquemment nues, généralement trimères (dimères chez
Gymnostachys), soit unisexuées et nues (
Arum). Si les fleurs sont unisexuées, elles sont généralement situées sur le même spadice, les fleurs mâles étant au-dessus des fleurs femelles. Le genre
Arisaema possède quelques espèces dioiques.
Quand ils existent, les 4-6 petits segments perianthaires peuvent être soudés et former une coupe. Les 1-12 étamines, libres ou soudées (
Colocasia), sont généralement extroses. Chez les genres unisexués, elles sont parfois remplacées par des staminodes dans les fleurs femelles. L'ovaire est supère ou semi-infère. Il comprend un ou plusieurs carpelles à une ou plusieurs loges, renfermant un ou plusieurs ovules anatropes, amphitropes ou orthotropes, sur des placentas basaux, apicaux, axiles (
Acorus) ou pariétaux. Le style unique a des formes variées, il peut être absent, et le stigmate est alors sessile. Ce dernier est hemisphèrique, capité ou discoide, exceptionellement lobé.
Le genre
Pistia, que Jussieu avait individualisé comme Pistiaceae, présente la particularité de posséder une spathe minuscule contennant deux étamines, équivalant morphologiquement à deux fleurs, soudées en une seule, surmontant un carpelle, équivalant à une fleur femelle. Cette inflorescence contractée, simulant une fleur, constitue un état préfloral qui évoque les
Lemnaceae.
L'inflorescence de certaines espèces dégage une odeur fétide écoeurante qui attire les mouches pollinisatrices. Parmi ces dernières, il a été montré que
Symplocarpus foetidus pouvait augmenter la température ambiante au sein de sa spathe, afi d'attirer encore plus ses vecteurs de pollinisation.
Arum dispose d'un mécanisme de pollinisation très évolué. La spathe enveloppant les fleurs est munie de poils dirigés vers le bas au-delà desquels les mouches atteignent le fond et y restent bloquées. Les mouches couvertes du pollen collecté sur les étamines d'une fleur mâle d'un autre pied d'
Arum, entrent en contact avec les fleurs femelles de la partie inférieure du spadice. Quand les poils du bas de la spathe se dessèchent, les mouches remontent vers le haut du spadice et se couvrent du pollen des fleurs mâles dont les étamines sont parvenues alors à maturité. Pour finir, les poils du haut de la spathe se dessèchent, permettant aux mouches de s'échapper et d'effectuer une pollinisation croisée en étant piégées dans une autre inflorescence.
Le fruit est une baie, parfois coriace, avec une ou plusieurs graines généralement albuminées avec un embryon droit, mais certaines espèces ont un embryon courbé avec ou sans albumen.


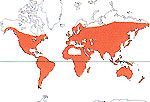 La famille est pantropicale, avec quelques espèces se trouvant en régions tempérées.
La famille est pantropicale, avec quelques espèces se trouvant en régions tempérées.























































































